|
STUDIES Department of Surgery II, Sahlgren's Hospital, Gothenburg University S-413 45 Gothenburg, Sweden Gothenburg 1987 Henrik Forssell: Studies on gastric bicarbonate secretion in man. Acta Chirurgica Scandinavica, Supplementum 540. 1987 HF Medical AB, Onsala, SWEDEN
ISBN 91-7900-2ISSN 0301-1860 STUDIES ON GASTRIC BICARBONATE SECRETION IN MAN
S-413 45 Gothenburg, Sweden
The investigations were performed in healthy subjects and duodenal ulcer (DU) patients, the latter group before and/or after proximal gastric vagotomy (PGV). Gastric bicarbonate secretion was determined by use of a gastric perfusion system in and computerized continuous pH and PCO2 Validation of the measuring system by small instilled amounts of bicarbonate gave a satisfactory correlation between added and recovered bicarbonate in the recorded range of bicarbonate determinations. Decreasing intragastric pH to between 3 and 4 did not affect the measured rate of bicarbonate secretion. Vagal stimulation increased gastric bicarbonate secretion in 16 healthy subjects from 410 ± 39 µmol/h to 692 ± 67 µmol/h (mean ± SEM, p <0.001). The muscarinic receptor antagonist, benzilonium bromide, almost abolished the sham feeding response while indomethacin left it nearly unchanged. Nine DU patients had identical basal and vagally stimulated bicarbonate output as healthy subjects. Basal bicarbonate secretion was significantly increased two months after PGV, whereas the bicarbonate output after sham feeding was unaltered. In the early postoperative period, anticholinergics reduced the enhanced basal bicarbonate secretion to a preoperative level. In six healthy subjects, graded fundic distension to volumes of 150 ml, 300 ml and 600 ml, each of 60 minutes, increased the bicarbonate secretion by 46 % (p < 0.05), 28 % (NS) and 84 % (p < 0.05), respectively. Continuous distension with 300 ml over 2.5 hours increased bicarbonate secretion, the peak response occurred at 45 minutes and gradually declined thereafter. Seven DU patients investigated after PGV had a response to graded fundic distension virtually identical to that in healthy subjects. Anticholinergics abolished the response to fundic distension, whereas indomethacin was without any significant effect. In healthy subjects, 16,16-dimethyl PgE2 gave an about threefold greater response than vagal stimulation, fundic distension or carbachol. It is concluded that gastric bicarbonate secretion in man is activated by cholinergic vagal nerves, the response being independent of intragastric pH above pH 2. There is probably an interplay of stimulatory and inhibitory mechanisms modulating basal and vagally stimulated gastric bicarbonate secretion. Fundic distension of the stomach stimulates bicarbonate secretion and this response is mediated by intramural neural cholinergic pathways. Vagal stimulation, fundic distension and carbachol appear to be sub maximal stimuli of bicarbonate secretion or else may have both stimulatory and inhibitory action on human gastric bicarbonate secretion. Key words: gastric bicarbonate secretion; gastric secretion; gastric; bicarbonate; humans; duodenal ulcer; vagal stimulation; fundic distension; prostaglandin; carbachol ISBN 91-7900-225-0 CONTENTS GASTRIC MUCOSAL DEFENCE MECHANISMS Introduction Pre-epithelial protection Mucus gel Bicarbonate secretion Epithelial protection Hydrophobicity of the epithelial lining Hydrogen ion permeation Sulphydryl compounds and EGF in gastric mucosal protection Gastric mucosal cell renewal Mucosal repair by restitution Subepithelial protection Mucosal blood flow Acid-base balance AIMS OF THE PRESENT INVESTIGATION MEASUREMENT OF HUMAN GASTRIC BICARBONATE SECRETIO The measuring system The measuring chamber The PCO2 electrode The computer recording Salivary bicarbonate Duodenogastric reflux of bicarbonate Experimental procedure and protocol The measuring system Basal bicarbonate secretion Vagal stimulation of gastric bicarbonate secretion Stimulation of gastric bicarbonate secretion by fundic distension Comparison between stimulants of gastric bicarbonate secretion This thesis is based on the following papers, which will be referred to inthe text by their Roman numerals: I Forssell H, Olbe L. Continuous computerized determination of gastric bicarbonate secretion in man. Scand J Gastroenterol 1985; 20:767-77 II Forssell H, Stenquist B, Olbe L. Vagal stimulation of human gastric bicarbonate secretion. Gastroenterology 1985; 89: 581-586 III Forssell H, Olbe L. Effect of proximal gastric vagotomy on basal andvagally stimulated gastric bicarbonate secretion in duodenal ulcer patients.Scand J Gastroenterol 1987;22:949-955 IV Forssell H, Olbe L. Effect of fundic distension on gastric bicarbonate secretion in man. Scand J Gastroenterol 1987;22:637-633 GASTRIC MUCOSAL DEFENCE MECHANISMS INTRODUCTION The stomach has a central role in the process of digestion. It is a reservoir and a mill and its glands secrete gastric juice containing hydrochloric acid and pepsin. Acidification of food destroys many ingested micro-organisms, begins the breakdown of food components and creates optimal conditions for protein digestion by pepsin. The stimulation of gastric secretion during digestion has been traditionally divided into cephalic, gastric and intestinal phases. During the cephalic phase, gastric secretion is evoked by stimulation of centres in the brain and impulses are mediated via vagus nerves. The gastric phase is initiated when food enters the stomach. The food volume stimulates stretch receptors in the gastric wall and food constituents interact with chemoreceptors resulting in further stimulation of acid secretion. The last stage, the intestinal phase, begins with the emptying of food and digestive products into the duodenum. This stage modulates both the secretion of acid and the emptying of food. The three phases overlap in time and are mutually interrelated in a complex nervous and humoral interplay. Gastric juice is produced by billions of cells located in the glands of the mucosa. The parietal cells, being one predominant cell type, have the ability to secrete a hydrochloric acid solution of pH 0.8 ie. the proton concentration of the primary gastric juice is about three million times greater than that of blood. The chief cells are another frequently occurring cell type in the glands and secrete pepsinogen, which is involved in the digestion of protein. Both acid and pepsin have proteolytic actions on living tissue and these components of gastric juice are a potential threat to the mucosa in terms of autodigestion. In the healthy stomach, there is a balance between aggressive factors and the protection provided by pre-epithelial, epithelial and subepithelial mechanisms of the mucosa. Secretion of mucus and bicarbonate by the surface epithelial cells constitute a mucus-bicarbonate barrier which is regarded to be a first line of defence. Other protective mechanisms comprise the inherent resistance of the epithelial cells to noxious stimuli as well as mechanisms for rapid tissue repair. The pathogenesis of peptic ulcer diseases multifactorial and far from established as yet. Apparently, the balance between aggressive factors and protective mechanisms in ulcer disease is upset in favour of ulcerogenesis. Possible mechanisms are excessive secretion of acid/pepsin, a reduced mucosal resistance or a combination of both. Superficial mucosal damage induced by e.g.ethanol oracetylsalicylic acid is usually followed by increased passive diffusion of luminal hydrogen ions into the mucosa. This increased permeability induced by weak acids and some other substances was first recognised by Teorell(1939) and the sequence of changes in the mucosa was further described by Davenport (1964, 1966). In the experiments by Davenport on canine Heidenhainpouches, injury was induced by a variety of agents. Damage was estimated in terms of increased flux of sodium and potassium into the gastric lumen and a loss of luminal hydrogen ions. Within the mucosa, the acid released histamine, which increased the permeability of the capillaries and further stimulated acid secretion. This resulted in interstitial oedema with haemorrhage and an additional breakdown of the mucosal barrier (Davenport 1967, Davenport 1970). Knowledge of gastric mucosal protection has increased rapidly during recent years. The following mechanisms have been suggested to be important and will be discussed in greater detail: pre-epithelial protection - mucus gel - bicarbonate secretion epithelial protection - hydrophobicity of luminal cell membranes - barrier to permeation - sulphydryl components - rapid cell turnover - restitution subepithelial protection - blood flow - tissue acid-base balance PRE-EPITHELIAL PROTECTION Mucus gel The entire surface of the gastric mucosa is covered by a continuous layer of mucus gel, which has a variable thickness of less than 500 µm (Bickel and Kauffman 1981, Kerss et al 1982, Allen et al 1983). Both the surface mucus cells and the mucus neck cells in the upper part of the glands secrete mucus. Mucus is released predominantly by the process of exocytosis (Zalewsky and Moody 1979). The main components of gastric mucus are neutral glycoproteins, which contain sialic acid and mucopolysaccharides. The glycoproteins, constituting the greater part of mucus, are of particular importance for their specific properties such as gel formation and viscosity. Native glycoproteins have a high molecular weight of about two million daltons and are formed by polymerization of four glycoprotein subunits, which are joined by disulphide bridges (Allen 1981). The subunit consists of a protein core with carbohydrate side chains. Each molecule of the porcine gastric mucus glycoprotein contains over 600 of these carbohydrate side chains. The most important sialic acid is N-acetylneuraminic acid. It is located in the terminal position of the carbohydrate side chain and has a strong negative charge. The rejection of neighbouring negatively charged groups of the glycoproteins results in molecular expansion and an increase in viscosity. The most important feature of the glycoproteins is, however, the hydrophilic action of the carbohydrate side chains. Water molecules are strongly attracted to the matrix with most of the molecules trapped within the interstices of the gel. Accordingly, about95 % of mucus consists of water. Mucus has several functions. It lubricates the mucosa and constitutes a first line of defence against noxious gastric contents. Of importance in the latter respect is the active secretion of bicarbonate from surface mucus cells. In addition, mucus retards diffusion of hydrogen ions, the rate in mucus being about four times slower than that in unstirred water (Williams and Turnberg 1980). Thus the mucus gel accomplishes a zone of limited mixing between the gastric luminal contents and the relatively small amount of secreted bicarbonate. The existence of a pH gradient across the mucus layer has been shown in vitro in the stomach of rabbits (Williams and Turnberg 1981), frog (Takeuchi et al 1983) and in vivo in the rat (Ross et al 1981). A pH gradient has also been demonstrated in the human stomach, in vitro (Bahari et al 1982) and in vivo (Quigley and Turnberg1985). The mucus gel has a very low permeability to large molecules such as pepsin, which should also be inactivated by the alkaline environment. However, pepsin continuously hydrolyzes the lumen-facing part of the mucus gel layer with production of mucus glycoprotein subunits. These have lost much of their viscous and gel-forming properties. The thickness of the mucus gel is thus determined by the dynamic balance between mucus secretion and surface erosions by proteolysis and mechanical destruction. The gel is resistant to hypertonic NaCl, bile, ethanol and indomethacin (Bell et al1985). Mucus secretion is stimulated, for example, by chemical irritants, carbachol, prostaglandins and the hormone secretin (Allen 1981). Another suggested property of mucus is facilitation of unidirectional flux of hydrogen ions from the gastric glands into the lumen. The transit of acid secreted by the parietal cells appears to occur through mucus which is more highly sulphated (Tasman-Jones 1985). Such mucus, with the highest negative charge contributed by sulphate and sialic acid radicals and produced especially by the mucus neck cells, should behave as a cation exchanger. A concentration gradient of sodium ions is proposed to be generated across the mucus layer by the continuous activity of a Na,K-ATPase at the basolateral membrane of the mucus cells. Sodium ions diffusing along this gradient should generate a diffusion potential positive at the cell-facing surface of the mucus gel. It is suggested that this potential moves hydrogen ions into the lumen and also prevents backwards diffusion (Smith et al1985). Gastric mucus is also important in the repair of superficial damage to the mucosa. After damage to the surface cells, there is a rapid release of copious amounts of mucus and plasma proteins which, together with cellular debris, form a continuous coat over the destroyed area (Wallace and Whittle1986). This mucoid cap provides a favourable micro-environment for repair by restitution. Bicarbonate secretion
Cyclic GMP, but not cyclic AMP, stimulates the alkaline secretion by gastric mucosa in vitro (Flemström 1977) and cholinergic stimulation increases the bicarbonate secretion in the canine antrum and fundus with a concomitant elevation in the mucosal concentration of cyclic GMP (Cheung and Newton 1979). This suggests that cyclic GMP may serve as an intracellular secondary messenger. For each hydrogen ion secreted by the parietal cell, a molecule of CO2 derived from arterial blood flow is converted to bicarbonate. The latter is released into gastric venous blood consequently provoking an alkaline tide after stimulation of acid secretion (Rune 1965).The vascular arrangement of the mucosa may facilitate the transport of the bicarbonate released by the parietal cells towards the bicarbonate secreting cells of the surface epithelium (Gannon et al 1984). Gastric as well as duodenal bicarbonate secretion has been extensively studied during the last 10 years both in vitro and in vivo preparations of various amphibia and common laboratory mammals, and especially by Flemström and his coworkers. It has been found that bicarbonate secretion from mucus cells is a metabolism dependent process rather than a passive diffusion and that it is stimulated and inhibited by a variety of physiological mechanisms and pharmacological agents (Flemström 1987). The rate of bicarbonate secretion from surface mucus cells is below 10 % of the maximum rate of acid secretion in several species (Garner et al 1983). Measurements of human gastric bicarbonate secretion have been accomplished by titration of bicarbonate concentration, from recordings of pH and PCO2 and from measurements of gastric juice osmolality. Gardham and Hobsley (1970) estimated basal bicarbonate secretion to be 236 ± 48 µmol/h (mean ± SEM) in patients with pernicious anaemia. The measurement was accomplished by means of back-titration of the alkaline gastric juice with hydrochloric acid. Andre and coworkers (1973) used a glycine buffer to trap hydrogen ions and bicarbonate secretion was subsequently determined with a titration technique in three steps, which included addition of HCl to the gastric sample to neutralize bicarbonate, removal of CO2 by boiling and finally back-titration to pH 7 with NaOH. They found the exceedingly high bicarbonate output of 7020 ± 840 µmol/h. It should be noted that the use of the glycine buffer caused an un physiological pH of over 9 in the stomach. Furthermore, such a buffer probably traps tissue CO2. In1982, Rees and coworkers reported a method for the measurement of human gastric bicarbonate secretion in which gastric juice was collected in 10-minintervals and pH and PCO2 were analyzed in the samples. This method was based on techniques developed for measurements of bicarbonate secretion in animals (Garner and Flemström 1978). The bicarbonate secretion was calculated using the Henderson - Hasselbalch's equation. Under basal conditions, it amounted to 347 ± 100 µmol/h. A similar method has been described by Johansson and collaborators (1983) who used a gastric instillation technique measuring pH and PCO2 in the recovered instillate. Kleist et al (1985) measured gastric bicarbonate concentration by using the van Slyke technique and found a gastric bicarbonate secretion value of similar magnitude as with the pH - PCO2 technique. Another approach for assessment of human gastric bicarbonate secretion has been described by Feldman (1983) using measurement of gastric juice osmolality. In healthy subjects, a basal gastric bicarbonate rate of 2600 ± 600 µmol/h was calculated. The method is based upon the fact that reaction between bicarbonate and hydrogen ions results in loss of osmoles and also on an assumed fixed relation between the osmolality of plasma and the osmolality of parietal and nonparietal secretions. Recently, both in vitro and human investigations have shown a several-fold overestimation by the osmolality method compared to both the pH- PCO2 method and back-titration (Odes et al 1987). Gastric bicarbonate secretion from amphibian mucosa and the
mucosa of various mammals is stimulated by humoral substances such as dibuturyl cyclicGMP
(Flemström 1977), calcium ions (Flemström and Garner 1980),cholecystokinin (Flemström
et al 1982) and several other gut hormones, for example, pancreatic glucagon, pancreatic
polypeptide and neurotensin (Konturek et al 1985). Gastrin and secretin have been found to
be without any stimulatory effect (Flemström 1987). Gastrin (Feldman 1983) and secretin
(Kleist et al 1985) were also unable to affect gastric bicarbonate output in man as was
the case with histamine (Feldman and Schiller 1982). Cholinergic agents, on the other
hand, stimulated bicarbonate secretion (Feldman 1983). Prostaglandins are potent
stimulants of gastric bicarbonate secretion (Garner and Heylings1979) and the synthetic
analogue of prostaglandin E2 has been shown to stimulate gastric bicarbonate secretion in
man (Johansson et al 1983, Fellowman 1983). Gastric bicarbonate secretion was increased by
electrical vagal stimulation in the cat (Fändriks and Delbro 1983) and by physiological
vagal stimuli such as sham feeding in man (Feldman 1985).Exposure to acid has been found
to increase gastric bicarbonate output in experiments with bullfrog mucosa (Heylings et al
1984), in the canine Heidenhain pouch (Konturek et al 1984), in the rat by acid solutions
of pH 1 but not pH 2 (Takeuchi et al 1986) and also recently in man when intragastric pH
was allowed to fall to 2 (Cramptonet al 1986). A vagally induced rise in feline gastric
bicarbonate secretion was enhanced by splanchnicotomy and/or ligation of the adrenal
glands (Fändriks 1986a), indicating adrenergic inhibition of the cephalic stimulated
gastric bicarbonate secretion. EPITHELIAL PROTECTION Hydrophobicity of the epithelial lining Both the surface mucus cells and the neck mucus cells face outermost towards the lumen of the stomach and constitute an anatomical barrier restricting passive diffusion into the underlying tissue of hydrogen ions and of sodium and potassium ions along their concentration gradients into the lumen. This tissue barrier may also prevent autodigestion by pepsin. Surfactants such as amphoteric phospholipids have the ability to increase the hydrophobicity of biological membranes and have been identified in gastric juice and on the apical cell membrane of the surface epithelial cells (Hills et al 1983, Butler et al 1983). The acid secreting part of the stomach in particular has a highly hydrophobic surface and these phospholipids have been suggested to resist digestion (Slomiany and Slomiany1980). The surfactant molecule is orientated in such a way that two hydrocarbon chains per molecule form a hydrophobic exterior surface. The opposite hydrophilic ends face the underlying cell, which is hydrophilic due to the preponderance of outward orientated hydroxyl and carboxyl groups in the membranes. The hydrophilic ends of the phospholipids are positively charged and form electrostatic bonds with the negatively charged cell membranes. Mucus makes a substantial contribution by stabilizing and replenishing the absorbed surfactant monolayer (Hills 1985). Surfactants contribute to the surface epithelial protection by preventing the water-soluble agents in the gastric lumen from reaching and damaging the cells. Exposure of the luminal surface of the stomach to a highconcentration of the damaging agents aspirin and sodium deoxycholate resultsin a rapid decrease in the hydrophobicity of the mucosa (Hills et al 1983). The reduction in surface hydrophobicity is effectively and dose-dependently reversed by synthetic analogues of prostaglandin E$FK022$FX(Lichtenberger et al 1985). Furthermore, a liposomal suspension of surface active phospholipids significantly protected the gastric mucosa of rats from experimental acid induced gastric damage(Lichtenberger et al 1983). Hydrogen ion permeation Hydrogen ion permeation across the epithelium may occur across the cell membranes or through tight junctions between the cells. The ion permeability of the tight junctions determines the electrical resistance of epithelia (Powel 1981). The resistance of the apical and basolateral cell membranes varies little between epithelia. The epithelium is moderately tight in the gastric fundus where as it is more leaky in the antrum. Permeation of hydrogen ions across tight junctions may occur by proton diffusion since protons can hop from one water molecule to another by rearrangement of hydrogen bonds. At neutral pH, the shunt paths are cation selective and relatively permeable to hydrogen ions. Protons and polyvalent cations such as Ca + neutralize the negative charges that line the pathways thereby changing the shunt path instead being anion selective. Thus, the negatively charged groups govern the permselectivity of the aqueos tight junctions. Consequently, at low pH the tissue may exclude hydrogen ion permeation (Powel 1981). Hydrogen ions may also be transported across the apical cell membrane by Na+K+ exchange controlled by intracellular calcium concentration (Benos 1982). When the mucosal surface epithelium is damaged, hydrogen ions diffuse from the lumen into the mucosa and sodium and potassium ions move from the tissue into the lumen. This increased permeability can also be observed as a marked decrease in the transmucosal electrical potential difference (PD). Acidification may damage the cells by interference with their ion transport mechanisms resulting in a loss of volume regulation and/or by denaturation of vital proteins.
Tissue ischaemia or injury produced by noxious agents such as ethanol may result in the accumulation of toxic free radicals. Ischaemia is followed by utilization of high energy compounds such as adenosine triphosphate and accumulation of adenosine monophosphate since oxidative phosphorylation is reduced. The adenosine monophosphate is catabolized further to hypoxanthine, which is accumulated (Younes et al 1984). Ischaemia also converts the enzyme xanthine dehydrogenaseto the xanthine oxidase form. This enzyme requires the presence of oxygen for activity. For instance after rapid reperfusion, oxygen becomes available for the tissue and xanthine oxidase will act on hypoxanthine producingsuperoxide radicals (O2), hydrogen peroxide and hydroxyl radicals. These free radicals, each containing an unpaired electron in its outer shell, are highly reactive, potent oxidizing and/or reducing agents. Cell injury is presumably caused by the damage of various cell membranes with intracellular release of e.g. lysosomal enzymes (Itoh and Guth 1985). Nonprotein sulphydryl compounds have been found in high concentrations in the gastric epithelium. The major component is reduced glutathione, which is capable of binding reactive free radicals. Ethanol lowers the concentration of nonprotein sulphydryls in the gastric mucosa whereas reduced glutathione induces gastric cytoprotection while increasing sulphydryl levels (Szabo et al 1981). Thus glutathione peroxidase appears to be a natural scavenger. Sulphydryl blocking agents prevent the cytoprotective action of certain prostaglandins concomitant with a decrease in sulphydryl concentration (Szabo et al 1981). Epidermal growth factor, EGF, which is a peptide of 53 amino acids, has been identified in salivary glands and in duodenal Brunner gland secretions. It is a powerful mitogen and stimulates DNA synthesis as well as ornithine decarboxylase activity. These observations suggest EGF to be important in the process of tissue repair and growth and in the physiological maintenance of the gastroduodenal mucosa. In addition, EGF is an inhibitor of gastric acid secretion. It has been found that EGF reduces gastric induced lesions at doses not inhibiting acid secretion (Skov Olsen et al1984). Approximately 10 % of the structure of EGF consists of SH-containing amino acids and it has been suggested that at least part of the protective effect of EGF in the gastroduodenal mucosa is due to these sulphydryl groups (Szabo 1984). Gastric mucosal cell renewal
Mucosal repair by restitution
SUBEPITHELIAL PROTECTION Mucosal blood flow The gastric mucosa, like all tissues, depends on blood flow for supply of nutrients and oxygen and for disposal of cellular waste products. Insufficient gastric blood flow is thus hazardous and a threat to tissue viability. When the gastric mucosa is exposed to an irritant that increases hydrogen ion permeation, tissue damage is greatly enhanced by a simultaneous decrease in mucosal blood flow. The exposure of canine gastric mucosa to sodium taurocholate and acid increased hydrogen ion diffusion and enhanced blood flow but occurred without gross damage to the mucosa (Ritchie 1975). However, simultaneous reduction in blood flow by vasopressin (Ritchie 1975) or by haemorrhagic shock (Ritchie and Shearburn1977) caused marked damage to the mucosa. Mucosal blood flow is thus essential in the disposal of hydrogen ions and noxious agents which permeate the mucosa and the ratio of blood flow to hydrogen ion diffusion determines the degree of the ensuing mucosal injury (Cheung and Chang 1977). Measurement of intramural pH of canine gastric mucosa in the absence or presence of luminal acid during haemorrhagic shock further suggested that the protective function of mucosal blood flow is related to the disposal of hydrogen ions entering the mucosa (Kivilaakso et al 1978a). Intramural pH decreased only slightly during reduction of mucosal blood flow when luminal acid was absent. However, a much greater decrease in the intramural pH occurred in the presence of luminal acid and a particularly rapid and profound decrease in intramural pH was observed after the addition of sodium taurocholate. Acid_base balance The intraglandular capillary supply of the stomach originates
from submucosal arterioles which divide into capillaries at the base of the glands. These
capillaries pass through the mucosa parallel to the glands, join to a capillary network
just beneath the surface mucus cells and drain into mucosal venules (Gannon et al 1984).
The mucosal venules do not receive further capillary tributaries deeper within the mucosa.
This vascular arrangement ensures the maintenance of an unidirectional blood flow in the
mucosal exchange vessels. Further, it may facilitate the protective role of alkaline tide
by transporting bicarbonate from lower regions of the mucosal vasculature to the
capillaries just below the surface mucus cells. In the oxyntic area of the stomach, the acid secretory process utilizes carbon dioxide. For each hydrogen ion secreted, one ion of bicarbonate is released. These bicarbonate ions diffuse into capillaries surrounding the glands and are transported by blood flow to the surface of the mucosa. Here, bicarbonate is delivered to mucus cells, but is also available for neutralization of hydrogen ions which may have diffused into the mucosa. The buffering capacity of tissue and blood, especially as related to the availability of bicarbonate ions to surface epithelial cells, is thus important in the protection of the gastric mucosa against acid induced injury. Actively secreting gastric mucosa with its accompanying alkaline tide of bicarbonate tolerates luminal acid far better than does a resting or inhibited mucosa (Kivilaakso et al 1978b, Arvidsson and Haglund 1984). In a rat model, parenteral administration of sodium bicarbonate
effectively protected the gastric mucosa from luminal acid (Kivilaakso 1981). This
protection depended on the presence of bicarbonate rather than on the alkalinity of the
tissue induced by respiratory hyperventilation. Nor did other buffer species offer any
protection. Ambient bicarbonate has also been shown to be important in the maintenance of
gastric mucosal intracellular pH (Kivilaakso 1983), which was significantly higher when
bicarbonate was present in the fluid surrounding isolated frog gastric mucosa than when it
was absent. Furthermore, intracellular pH was higher than extracellular pH only when
bicarbonate was present. Consequently, nutrient (blood) bicarbonate ions may enhance the
ability of the mucosal cells to withstand acid. The ability of the gastric mucosa to resist autodigestion is of multifactorial nature. Both mucus and bicarbonate are actively secreted by the mucus cells of the gastric mucosa and constitute a mucus-bicarbonate barrier, which is regarded as a first line of defence. While gastric bicarbonate secretion has been extensively characterized in vitro and in vivo in several mammals, the nature of this bicarbonate secretion has been very little explored in humans. The aims of the thesis were
MEASUREMENT OF HUMAN GASTRIC BICARBONATE SECRETION Measurement of gastric bicarbonate secretion in the present study was based on the assumption that bicarbonate appearing in the gastric perfusate reflected the secretion of bicarbonate into the mucus gel layer. Acid inhibition by histamine H2 -receptorantagonists increased intragastric pH to a level above pH 6. At pH 6.10, i.e. at the pKa of carbonic acid, half of intragastric bicarbonate exists in the form of free bicarbonate ions, the rest being converted to carbon dioxide by the reaction of bicarbonate with acid. Consequently, use of a higher intragastric pH of around 6.5 further decreased the formation of carbon dioxide and thus minimized the possible loss of CO2 by eructation, passage through the pylorus or diffusion into the gastric wall. The measuring system contained electrodes which simultaneously measured pH and PCO2. The concentration of free bicarbonate was calculated according to the formula of Henderson (1908) and Hasselbalch (1916). A proportion of free bicarbonate was always neutralized by hydrochloric acid with formation of CO2. The t˝ for this reaction is about 14 seconds (Flemström and Garner 1987). The concentration of CO2 was added to obtain total bicarbonate concentration according to the formulae:
The solubility coefficient, S, for CO2 and the dissociation constant, pKa, for carbonic acid, both depend on the solution and its chemical and physical properties. For example, the solubility coefficient increases with decreasing temperature. At 37°C, 0.033 mmol per mm Hg of CO2 is dissolved in 1 litre of water. The corresponding values at 30°C and 20°C are 0.039 and 0.052 mmol per mm Hg, respectively. Presence of ions in the solution depresses the S value (van Slyke et al 1928), especially at ion concentrations above 0.3 M. The dissociation constant, pKa, will increase slightly with decreasing temperature but is affected more by the ionic strength of the actual solution. Hence, the pKa of carbonic acid is reduced by 0.5 x the square root of the ionic strength of the solution (Hastings et al 1925). The pKa in water solutions at 37°C is 6.31 and the corresponding value in plasma is 6.10. The measuring system A Salem sump tube no. 12 was positioned with its tip in the
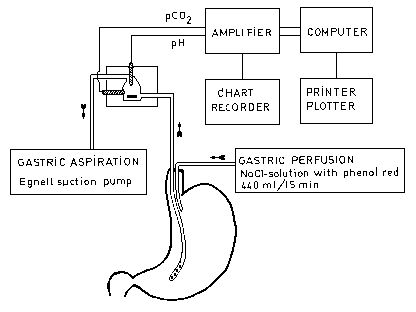
upper part of the gastric antrum and was coupled to a measuring system. Its outlet was
connected to an Egnell suction pump, which produced an intermittent negative pressure of
15 - 20 kPa once a second (Fig. 1, I).
The concentration of phenol red in the perfusate was 8 mg/l. Concentration of phenol red in the aspirate was measured spectrophotometrically at a wavelength of 560 nm. A high perfusion rate of 440 ml/15 min facilitated mixing of secreted gastric juice with the perfusate. In addition, the high rate diluted secreted acid and the rapid washing-out shortened and allowed easy detection of episodes of duodenogastric reflux of bicarbonate. The measuring chamber The measuring chamber was made from translucent plastic material and occupied a volume of only 9 ml (I). The pH and P CO2 electrodes were placed in such a manner that air bubbles did not interfere with the recordings. The outlet of the measuring chamber was at the top allowing air bubbles which were intermingled with the gastric juice to pass along the side wall to the top of the chamber without disturbing measurements of pH and P. The pH electrode and its reference (Radiometer, Denmark: G2040C and K8040 respectively) both had screw-caps allowing gas-tight insertions into the measuring chamber. A magnetic stirrer inside the measuring device ensured thorough mixing of the passing gastric aspirate. The P CO2 electrode The PCO2 electrode(Radiometer, Denmark: E8001) is basically a pH electrode placed in bicarbonate electrolyte behind a teflon membrane of 12 µm thickness. This membrane is impermeable to ions such as HCO3 and H+ but permeable to gas molecules. The CO2 dissolved in the gastric perfusate diffuses into the bicarbonate electrolyte until an equilibrium is reached, changing its pH. This pH change is converted into a P CO2 reading in the amplifier (Radiometer, Denmark: PHM 73) by using a linear calibration curve which relates log PCO2 to pH. The PCO2electrode was always calibrated against two carefully analyzed gas mixtures which contained CO2 in the range of 1 to 3 % in N2 for alternate 15-min periods in the few hours before the experiment. The computer recording The pH and PCO2 signals from the amplifier were stored digitally in a computer (Commodore, USA). The amplifier was provided with an analog-digital converter which was interfaced between the output of the amplifier and the computer. The binary pH and PCO2 signals were preceded by identification signals to enable the computer programme to differentiate between pH and PCO2 signals. The computer calculated total bicarbonate concentration every 30 seconds. Since the PCO2 electrode is rather slow-reacting compared with the pH electrode, the computer programme compensated in the calculations by using the pH value recorded 90 seconds before the PCO2 value. The computer was programmed to use a pKa = 6.10 and a S value = 0.039 in the calculation of total bicarbonate concentration since the aspirate had a temperature of about 30°C. During measurement of gastric bicarbonate secretion the computer stored 252 recordings and performed 256 calculations per hour. Salivary bicarbonate The salivary concentration of bicarbonate varies from a few mmoles/litre to about 25 mmoles/litre. The subjects were instructed not to swallow their saliva and it was collected with the aid of a dental suction set. The volume of saliva collected during unstimulated conditions was up to 20 ml per 15 minutes. Even when saliva was expectorated, the involuntarily swallowed salivary bicarbonate amounted maximally to 13 % of the measured bicarbonate in gastric contents (Forssell et al 1984). The amount of involuntarily swallowed saliva was estimated in the present investigation by measuring the concentrations of amylase in the saliva and the 15-min gastric aspirates:
Any contribution to amylase by duodenogastric reflux was assumed
to be negligible as long as bilirubin was absent from the recovered gastric perfusate
and/or any rapid transient increases in bicarbonate concentration from duodenogastric
reflux episodes (see below) did not occur. The total salivary bicarbonate concentration
was determined from measurement of salivary pH and PCO2 as described for
gastric samples. The amount of swallowed salivary bicarbonate was calculated from the
volume of swallowed saliva and its bicarbonate concentration:
Finally, the amount of bicarbonate in swallowed saliva was subtracted from the calculated total gastric bicarbonate output to obtain the amount of bicarbonate secreted by the stomach:
Duodenogastric reflux of bicarbonate The occurrence of alkaline duodenogastric reflux was detected from bile staining of the gastric perfusate passing through the transparent measuring device and tubes. Coincident with the visual observation of bile reflux was a short rise in recorded pH and PCO2 values (I). Such spikes were superimposed on the basal level of bicarbonate secretion and were usually of only a few minutes duration. These spikes were excluded in the calculations of gastric bicarbonate output by a computer-editing programme. Moreover, measurement of bilirubin concentration at alkaline pH in the collected 15-min gastric aspirates excluded continuous modest duodenogastric reflux. Periods with a bilirubin concentration > 1 µmol/l were rejected. Experimental procedure and protocol Bicarbonate secretion was measured after an overnight fast. Before the experiment, a histamine H2-receptor antagonist was given orally to the subjects. Two alternative premedication regimens were used in the experiments, either cimetidine, 400 mg, one and three hours before the investigation or ranitidine, 150 mg, two and fourteen hours before start of the experiment. Both regimens gave an increase in intragastric pH to around 6, which usually lasted for about four hours. The pH electrode was calibrated at pH 6.00 and 7.00 with two buffer solutions. Thereafter, the PCO2 electrode was calibrated against two different concentrations of CO2 for alternate 15-min periods. The subject was seated in a semi recumbent position and the Salem sump tube with the additional small tube for perfusion was introduced into the distal part of the stomach. Residual gastric juice was aspirated and discarded. The perfusion system and the computer based system for measurement of gastric bicarbonate concentration were subsequently commenced. Aliquots of fifteen minutes aspirate were used for marker determination and for measurement of amylase and bilirubin concentrations. The gastric bicarbonate output values and the amount of swallowed saliva were stored in a computer database (Digital Equipment Corp., USA). Statistical analyses were performed with the aid of an RS/1
computer programme (Bolt Beranek and Newman Inc., USA). Results are expressed as mean ±
SEM. Statistical significance was evaluated using Wilcoxon's matched-pairs signed rank
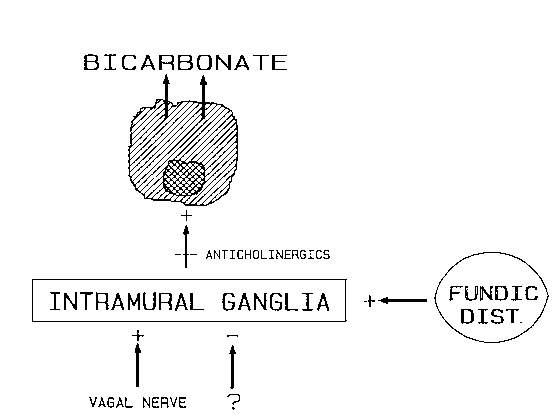
test. The measuring system In ex vivo control experiments, acid was added to bicarbonate solutions in the chamber of the measuring device. The time required to receive stable recordings of PCO2 varied from 0.5 to 3 minutes, depending on the amount of CO2 formed during the process of neutralization. In contrast, stable recordings of pH were obtained almost immediately. Thus, the computer had to compensate for the slower reaction of the PCO2 electrode by using the pH value recorded 90 seconds before that of PCO2. In other ex vivo experiments, the interference of air bubbles in the perfusate was tested. The recordings of pH and PCO2 were disturbed only when large amounts of air bubbles occurred in the effluent. A high rate of gastric perfusion with few air bubbles from the air inlet of the Salem sump tube would thus provide more reliable measurements in human experiments. Two perfusion rates were therefore tested in seven healthy subjects. Rates of 220 ml/15 min, which is the rate routinely used in our acid secretory studies, and 440 ml/15 min were evaluated. Less variation in the recorded basal secretion of bicarbonate was observed with the higher perfusion rate (I). This may reflect the reduced number of air bubbles contained in the effluent. The higher perfusion rate of 440 ml/15 min was thus used in the subsequent studies of gastric bicarbonate secretion. To further validate the measuring system, exogenous bicarbonate in amounts varying from 50 to 400 µmoles was instilled into the stomach of healthy subjects. The correlation coefficient, r, between added and recovered amounts of bicarbonate was 0.91 (p < 0.001, I). This indicates that the bicarbonate output was satisfactorily measured over a wide range of secretory rates. With a luminal pH at the pKa of carbonic acid, 50 % of bicarbonate occurs as free ions and the rest reacts with acid to form CO2. At values of intragastric pH below 4, virtually all bicarbonate is in the form of CO2. The effect of decreasing intragastric pH on recorded values of bicarbonate output was studied in healthy subjects by adding 250 µmol HCl to the perfusate during one 15-min period. The intragastric pH fell to between 3 and 4 but no change in bicarbonate output was observed during the 15-minperiod of acid load or in subsequent 15-min periods (I). It could thus be concluded that the system adequately measured bicarbonate even if the intragastric pH fell to between 3 and 4 and that a moderately low intragastric pH did not stimulate bicarbonate secretion. Similar validation experiments were performed by Johansson and coworkers (1983) when testing their technique using gastric instillations. These investigators were unable to demonstrate a stimulation of bicarbonate secretion by a pH of about 3.Recently, Crampton et al (1986) reported the stimulatory effect of an intragastric pH of about 2 on gastric bicarbonate output in man. In animal experiments, stimulation of bicarbonate secretion by luminal acid occurs both in the stomach and the duodenum, but the required acidity of the gastric solution needs to be about pH 2 (Heylings et al1984) or even lower (Takeuchi et al 1986). The present results also indicate that loss of CO2 by eructation, passage through the pylorus or diffusion into the gastric tissue is small and presumably negligible. Similar findings have been made by Fändriks in cats (1986b). He reported a modest underestimation of bicarbonate secretion during active acid secretion. Moreover, animal experiments have shown that diffusion of CO2 out of the stomach as well as diffusion of CO2 into the stomach from tissue and blood is small (Garner and Flemström 1978, Kauffman and Steinbach 1981). Hence, the findings of a high intragastric PCO2 value, in the present investigations sometimes above 40 mm Hg and thus exceeding that in blood, and even higher values reported by others (Schierbeck 1892, Rune and Henriksen 1969, Garner and Flemström 1978) indicate that CO2 is produced in the gastric lumen by neutralization of bicarbonate with acid. To minimize the risk for loss of CO2 in the present experiments, inhibition of gastric acid secretion by histamine H2-receptor antagonists was used, particularly since cimetidine has been found to exert no direct effect on bicarbonate secretion in animals (Garner and Flemström 1978, Flemström and Turnberg 1984). It should be mentioned, however, that a high intragastric pH may to some extent decrease gastric bicarbonate secretion by removing the stimulatory effects of luminal hydrogen ions, as discussed above. The measuring system enables detection of alkaline duodenogastric reflux episodes as indicated by a short rise in both pH and PCO2. These spikes frequently coincided with the visual observation of bile reflux and were superimposed on the bicarbonate secretion level. The short duration of these reflux-generated bicarbonate spikes of only a few minutes very probably reflects the use of a high rate of gastric perfusion to wash-out alkaline reflux. Moreover, measurement of bilirubin concentration in the collected gastric samples excluded every continuous modest duodenogastric reflux. In a series of experiments in sixteen healthy subjects (mean age 30 years, range 20-46 years, II), no duodenogastric reflux was observed at all in 5 subjects. Eleven subjects had a mean of 4.6 episodes (range 1 - 12) over 105 min of measurement of basal bicarbonate secretion. Less than half of these episodes occurred without simultaneous bile staining of the aspirate. These results indicate that the present method distinctly demonstrates duodenogastric reflux, which thus can be eliminated enabling correct calculation of gastric bicarbonate secretion. In summary, the various validation tests have shown that the computerized measurement of human gastric bicarbonate secretion satisfactorily determines bicarbonate output. Continuous measurement of pH and PCO2 seems preferable to discontinuous recording of these parameters. Moreover, the method detects duodenogastric reflux without intubation of the duodenum. A high gastric perfusion rate facilitates the identification of duodenogastric reflux episodes and shortens their duration by rapid washing-out. Basal bicarbonate secretion The rate of basal bicarbonate secretion in 34 healthy subjects (10 women and 24 men) with a mean age of 32 years (range 20 - 46 years) amounted to 366 ± 23 µmol/h (mean ± SEM). These subjects have been investigated on a total of 107 different occasions and the intra-individual coefficient of variation was 30 %, a value much less than the 80 % (Feldman and Richardson 1981) and 110 % (Lind et al 1986) reported from studies on human basal acid secretion. The larger variation observed in basal acid secretion may be attributed to the influence of a varying vagal tone affecting the parietal cell mass. Basalacid secretion can be inhibited by anticholinergics or vagotomy (Gillespie et al 1960, Stenquist et al 1979). In contrast, a rather high dose of anticholinergics (benzilonium bromide) was unable to change basal gastric bicarbonate secretion in healthy subjects (II). This may suggest that any cholinergic drive at the mucus cell level does not modulate basal gastric bicarbonate secretion in the intact human stomach. The cyclooxygenase inhibitor, indomethacin, did not significantly affect basal gastric bicarbonate secretion (II), indicating that basal gastric bicarbonate secretion does not depend on the release of endogenous prostaglandins. This is in contrast to the findings in the human duodenum (Selling et al 1987) where indomethacin reduced both basal and acid stimulated bicarbonate secretion. The rate of basal gastric bicarbonate secretion in healthy subjects as determined in the present investigations agrees with the findings by other laboratories using recordings of pH and PCO2 to calculate the bicarbonate secretion (Rees et al 1982, Johansson et al 1983). Much higher rates of basal bicarbonate secretion in humans, about 2500 µmol/h, however, have been reported by Feldman (1983). Measurements of .gastric volume, hydrogen ion concentration and osmolality of both gastric juice and plasma were used to calculate secretion by the method of Feldman. The calculation is based on a two component model of gastric secretion and the assumption of a fixed relation between the osmolalities of plasma and parietal and nonparietal secretions. However, this relation may not be constant and, moreover, has been examined only in the stimulated canine stomach. Another explanation for the quantitative discrepancy in rates of basal gastric bicarbonate output is that hypotonicity of gastric juice could arise from both the neutralization of secreted bicarbonate by acid and the processes for secretion of acid and water within the gastric glands (Flemström 1985). Although it seems that the osmolality method overestimates secretion of human gastric bicarbonate (Odes et al 1987), results based on osmolality and pH - P CO2 measurements, respectively, seem to be qualitatively similar. The basal bicarbonate secretion rate in nine patients with duodenal ulcer disease (mean age 52 years, range 33 - 69 years) was 414 ± 57 µmol/h (III) and did not differ significantly from that in healthy subjects. Basal gastric bicarbonate secretion was increased by 30 % to 539 ± 74 µmol/h (N =9, p < 0.01) about 2 months after proximal gastric vagotomy (III). Hypothetically, the effect of partial denervation of the stomach by proximal gastric vagotomy may be consistent with removal of an inhibitory mechanism modulating basal gastric bicarbonate secretion. In support of this hypothesis, benzilonium bromide restored the enhanced basal bicarbonate secretion after vagotomy to a preoperative level (III). Benzilonium bromide had no effect on basal gastric bicarbonate secretion in the intact stomach(II), possibly due to an active inhibitory mechanism. The enhanced basal gastric bicarbonate secretion must originate either from the innervated and/or denervated region of the stomach. However, the present investigation was unable to further clarify which area of the stomach that had increased its bicarbonate secretion. The increase may originate from the antrum in analogy with the regulation of canine antral secretion of gastrin (Debas et al 1975) where a vagally dependent oxyntopyloric reflex inhibition exists. The increased gastric bicarbonate secretion may thus be explained by an enhanced secretion from the antrum caused by removal of such a hypothetical inhibitory mechanism following proximal gastric vagotomy. The antrum accounts for about 25 % of the total surface area of the stomach (Ito 1981) and evidence has been presented (Flemström 1977, Konturek et al 1985) that the bicarbonate secretion in the antrum and the fundus-corpus region of the intact stomach is similar. Assuming that the basal gastric bicarbonate secretion from the vagally denervated fundus and corpus area is unchanged after proximal gastric vagotomy, the increase in basal bicarbonate rate from the antrum would be about 120 %. This figure would correspond well with the increase in gastric bicarbonate secretion, about 125 %, two months after vagotomy in response to sham feeding (III). It is reasonable to presume that this response originates only from the antrum. In the conceptual model of gastric bicarbonate secretion (Fig.
2), one of the final transmitters close to the mucus cell level most probably is
acetylcholine. The nature of the inhibitory mechanism likely to affect the intramural
ganglia is not clear. It may be mediated via noncholinergic transmission since a rather
high dose of benzilonium bromide did not affect the basal bicarbonate secretion of healthy
subjects with intact vagal nerves. The influence on gastric bicarbonate secretion of
noncholinergic vagal fibres would accord with a previous demonstration in the cat, in
which the transmitter was suggested to be substance P (Fändriks and Delbro 1983).
Moreover, a noncholinergic vagal release of gastrin and a cholinergic inhibitory mechanism
controlling gastrin release have been shown in man (Feldman et al 1979, Stenquist et al
1979). A vagal cholinergic stimulatory effect on the gastric motor activity and a vagal
noncholinergic, probably VIP-ergic, relaxatory effect on the stomach have also been
demonstrated (Martinson and Muren 1963, Fahrenkrug et al 1978). The inhibitory mechanism
may consist of vagally dependent oxyntopyloric reflexes, as discussed above, (Debas et al
1975)which are eliminated by proximal gastric vagotomy. Fig. 2 Conceptual model of the regulation of human gastric
bicarbonate secretion. Nevertheless and irrespective of what the nature of this inhibitory mechanism may be, the basal bicarbonate secretion had declined and was at the preoperative level about one year after the proximal gastric vagotomy, suggesting a gradual regress of the non-inhibited basal gastric bicarbonate secretion. Vagal stimulation of gastric bicarbonate secretion Sham feeding is a physiological, vagally mediated stimulus of gastric acid secretion in man (Stenquistet al 1978). Modified sham feeding is accomplished by allowing the subject to chew and spit out a meal (Noring 1951). The acid response to modified sham feeding is about 50 % of the maximal acid response to pentagastrin. It is of a similar magnitude to that produced after adequate sham feeding, determined in the postoperative period in patients in whom food was swallowed and subsequently drained via a gastrostomy avoiding chemical stimulation of the stomach by food (Stenquist et al1978). The acid response to sham feeding can be inhibited by about 65 % but not abolished by anticholinergics, suggesting that the neurotransmission at acid-secreting cells is only in part cholinergic and that peptidergic transmission or amines may also be involved (Stenquist et al1979, Stenquist et al 1987). Vagal stimulation by sham feeding, performed identically as in
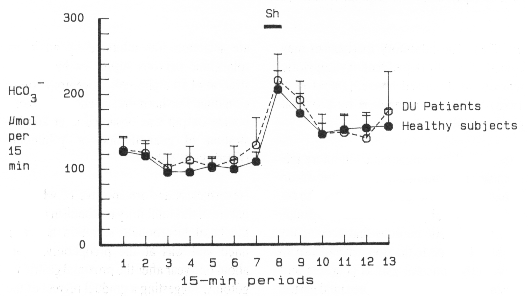
acid secretory studies, was used in the present study to examine whether stimulation of
the surface mucus cells increased bicarbonate secretion. Vagal stimulation in sixteen
healthy subjects increased gastric bicarbonate output by 69 %, i.e. from 410 ± 39 to 692
± 67 µmol/h (mean ± SEM, p < 0.001, II). This increase was mainly caused by an
increase in bicarbonate concentration, and to a lesser extent, by an increase in volume.
In most experiments, the gastric bicarbonate response to sham feeding began only a few
minutes after start of the vagal stimulation with a peak bicarbonate output in the 15-min
period of sham feeding (Fig. 3). In cats, electrical stimulation of vagal trunks induced a
rapid increase in bicarbonate output. This rise in bicarbonate secretion occurred somewhat
earlier than that of hydrogen ion secretion, suggesting anticipatory protection by
bicarbonate against acid (Nylander et al 1987).In the present investigation, vagal
stimulation evoked a similar increase in gastric bicarbonate output irrespective of
intragastric pH, range above 2 to about 7 (II). This indicates that the bicarbonate
response during sham feeding is not secondary to a concomitant increase in luminal
hydrogen ion concentration. The anticholinergic drug, benzilonium bromide, which has a
minimal ability to cross the blood-brain barrier, inhibited the rise in gastric
bicarbonate secretion in response to sham feeding by 88 % (II). The bicarbonate response
to vagal stimulation was thus almost abolished by benzilonium bromide, suggesting a mainly
cholinergic excitatory effect by sham feeding on human gastric bicarbonate secretion. The vagal stimulation of gastric bicarbonate output in nine duodenal ulcer patients did not differ significantly to that of healthy subjects (Fig. 3), the increase being 67 % above basal bicarbonate secretion i.e. from 414 ± 57 to 691 ± 83 µmol/h (p < 0.01, III). After proximal gastric vagotomy, which was complete in all patients since no increase in acid output was observed after a separate sham feeding test, there was still a significant increase (p < 0.01) in gastric bicarbonate secretion, which amounted to 29 %, in response to vagal stimulation. Presumably, this response originated from the still vagally innervated antral part of the stomach. The bicarbonate output, however, is about twice that expected from the antrum. This seems consistent with the observation of an increased secretion also during basal conditions and may be due to the loss of an antral inhibitory mechanism. A third sham feeding test was performed about one year after proximal gastric vagotomy. The response to vagal stimulation was then abolished by anticholinergics which implies a remaining susceptibility to anticholinergic blockade (III). Quantitatively, the human gastric bicarbonate secretory response to vagal stimulation is much smaller than the acid response to sham feeding. This indicates, on allowing for the neutralizing effect of the secreted bicarbonate, that the amount of acid produced in conjunction with sham feeding clearly is still below the maximal acid secretory capacity determined by a pentagastrin test. Consequently, this implies that sham feeding in man is either a submaximal stimulus of acid secretion or affects both stimulatory and inhibitory acid secretory mechanisms. The gastric bicarbonate response to sham feeding may also be submaximal or sham feeding may affect both stimulatory and inhibitory mechanisms. A much greater response, about a 200 % increase, was thus observed after instillation of 16,16 dimethyl prostaglandin E2 (I, Johansson et al1983). The occurrence of inhibitory actions would be consistent with findings in the cat and rat of a sympathetic reflex inhibition of vagally mediated gastric and duodenal bicarbonate secretion (Fändriks 1986b, Jönson and Fändriks 1986). Stimulation of gastric bicarbonate secretion by fundic distension The second phase of gastric acid secretion is the gastric phase, initiated by food reaching the stomach. Both chemical and mechanical stimuli are involved in this process. Graded distension by stepwise inflation of a balloon located in the fundic area of the stomach elicits a volume-related acid secretory response (Grötzingeret al 1977a). This acid response to fundic distension was similar both in healthy subjects and duodenal ulcer patients and the observed peak acid response averaged about 50 % of the peak in response to pentagastrin stimulation. Graded fundic distension to volumes of 150 ml, 300ml and 600 ml increased the acid output in six healthy subjects by 94 %, 106% and 194 %, respectively (Grötzinger et al 1977a). Experiments with graded fundic distension of the stomach in six
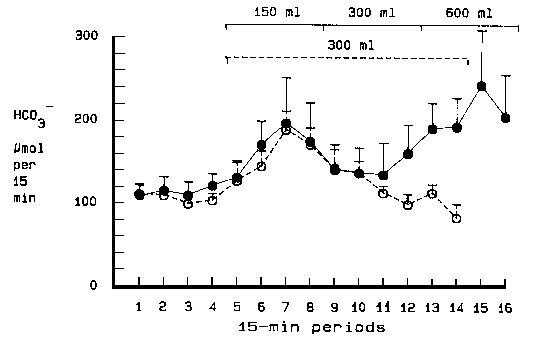
healthy subjects evoked an increase in gastric bicarbonate output amounting to 46 %(p <
0.05), 28 % (NS) and 84 % (p < 0.05) over 60 minutes of distension to volumes of 150
ml, 300 ml and 600 ml, respectively (Fig. 4). Obviously, no dose-dependent bicarbonate
secretion was observed with this experimental design, which was identical with that used
in the acid secretory experiments by Grötzinger and coworkers (1977a). Instead, a peak in
bicarbonate secretion occurred after 45 minutes of distension with 150 ml, followed by a
decline. During distension to a volume of 300 ml, bicarbonate secretion was lower than in
the preceding period with 150 ml and not significantly different from basal secretion. A
second peak occurred 45 minutes after start of distension with 600 ml. A series of
experiments were performed in four of the six healthy subjects to further clarify the
nature of this biphasic response to graded fundic distension. Distension with a volume of
300 ml was maintained for 150 minutes. Again, there was a peak after 45minutes.
Bicarbonate secretion then gradually declined and reached basal bicarbonate levels 60
minutes later despite the continued distension (Fig.4). These results indicate that fundic
distension elicits a fading bicarbonate response, possibly reflecting volume adaptation of
the gastric wall. Another possibility to explain the decrease in bicarbonate output during
sustained fundic distension would be a somewhat slower activation of inhibitory
mechanisms. In fact, Schöön et al (1978) have shown that distension-induced inhibitory
mechanisms operate in the regulation of gastric acid secretion. Moreover, experiments in
healthy subjects as well as in vagotomized duodenal ulcer patients with fundic distension
failed to enhance pentagastrin-induced acid secretion. Instead the distension inhibited
acid output in some subjects (Grötzinger et al 1977b). In seven duodenal ulcer patients, who previously had undergone a proximal gastric vagotomy, graded fundic distension resulted in virtually an identical biphasic response as in healthy subjects (IV). All patients were considered to be completely vagotomized, since postoperative tests showed no acid increase in response to sham feeding. On the reasonable assumption that the two groups are comparable, it seems that the distension-induced bicarbonate response is mediated mainly by short enteric intramural neural pathways and not by long vagovagal reflexes (Fig. 2). It was also observed that the fading response to graded distension remained after proximal gastric vagotomy. The response to fundic distension with 150 ml for 30 minutes was abolished in healthy subjects by pretreatment with the muscarinic antagonist, benzilonium bromide, while pretreatment with the cyclooxygenase inhibitor, indomethacin, was without any effect (IV). These results would suggest a purely cholinergic reflex mechanism which is not mediated by endogenous prostaglandin liberation. The latter accords with findings in earlier studies of gastric bicarbonate secretion in man (II, Feldman and Colturi1984) but contrasts results from studies of human duodenal mucosal bicarbonate secretion (Selling et al 1987). In the latter study, indomethacin was given orally in the same dose as in the present investigation, but was shown to reduce both basal and acid-stimulated duodenal bicarbonate secretion. Comparison between stimulants of gastric bicarbonate secretion In the experiments reported (I,II,IV), bicarbonate secretion in
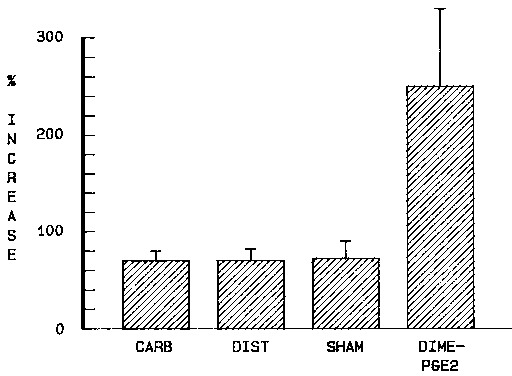
four of the healthy subjects had been stimulated by sham feeding, as well as by fundic
distension and by prostaglandin administration. An additional series of experiments were
performed in these subjects. The effects of increasing doses of carbachol in the range of
100 to 200 µg subcutaneously were tested. The purpose was to compare their maximal
responses to the different stimuli. The maximum dose of carbachol tolerated was 200 µg,
which caused a slight bradycardia, an increase in salivary concentration of amylase and
some abdominal motor discomfort. Sham feeding for 15 minutes, fundic distension to a
volume of 600 ml and 150 - 200 µg of carbachol all increased bicarbonate output by about
70 % above basal level. Instillation of 16,16-dimethyl prostaglandinE2, 80 µg for 5
minutes, gave an about threefold greater response (Fig. 5). These findings suggest that
vagal stimulation, fundic distension and carbachol are submaximal stimuli of human gastric
bicarbonate secretion that they have both stimulatory and inhibitory actions on
bicarbonate secretion. Fig. 5 Increase in gastric bicarbonate secretion in four healthy
subjects in response to carbachol 150-200 µg subcutaneously (CARB), fundic distensionto a
volume of 600 ml (DIST), 15 minutes of sham feeding (SHAM) and gastricinstillation of 80
ßµßg 16,16-dimethyl prostaglandin E2 for 5 minutes (DIME-PGE2). Per cent increase above
basal level. Mean + SEM. The findings in the present investigation suggest a complex interplay between stimulatory and inhibitory mechanisms influenzing basal as well as stimulated human gastric bicarbonate secretion. Further measurements of human gastric bicarbonate secretion in more defined subgroups of subjects or patients may improve the possibility of clarifying these mechanisms. The complexity of the regulation of bicarbonate secretion is highlighted by the vagally dependent stimulatory effect of the muscarinic M2-antagonist, pirenzepine, on mucosal bicarbonate secretion in the rat duodenum (Säfsten and Flemström 1986). The role of a sympathetic nervous influence on gastric bicarbonate secretion is of considerable interest and calls for further investigation. Such a sympathetic nervous control of vagally stimulated gastroduodenal bicarbonate secretion has been shown to exist in cats (Fändriks 1986b). Current experiments in healthy subjects in our laboratory also suggest a sympathetic tone to regulate basal gastric bicarbonate secretion. Since the pathogenesis of peptic ulcer disease is multifactorial and the disease, in many respects, is of psychosomatic nature, an enhanced sympathetic drive associated with mental and physical stress may contribute to a decreased epithelial bicarbonate secretion and result in a defective mucus-bicarbonate barrier. Recently, it has been shown that duodenal ulcer patients may have an impaired proximal duodenal mucosal bicarbonate secretion (Isenberg et al 1987). Since duodenal ulcer patients in the present investigation had a normal gastric bicarbonate secretion during basal or vagally stimulated conditions, the findings may indicate that ulcerogenesis is caused by local pathological factors in the duodenum and be due to an increase in aggressive factors combined with a decrease in local mucosal resistance. Patients with prepyloric gastric ulcers have acid secretory values similar or slightly lower than those of duodenal ulcer patients but heal their ulcers very slowly and their treatment is often a considerable challenge to both the physician and surgeon. On the other hand, in the pathogenesis of gastric ulcer disease, acid and pepsin secretion are usually within normal limits or reduced. Hence, a defect in the local gastric mucosal defence mechanism is suggested to operate in the development of both prepyloric and gastric ulcers, prompting the need for further investigations of gastric bicarbonate secretion in these patients. If such a defect in gastric bicarbonate secretion can be revealed, therapy may also be directed to the development of agents that enhance gastric bicarbonate secretion and thus strengthen the protective mucus-bicarbonate barrier.
The investigations were performed in healthy subjects and duodenal ulcer patients, the latter group before and/or after a proximal gastric vagotomy operation. Healthy subjects as well as ulcer patients were premedicated with a histamine H2-receptor antagonist and gastric bicarbonate secretion was determined by use of a gastric perfusion system in combination with computerized continuous recordings of pH and PCO2. The contribution of alkaline saliva to the measured gastric bicarbonate secretion was minimized by continuous salivary suction and correction was made for swallowed saliva by measurement of amylase in the gastric aspirate. A high rate of gastric perfusion facilitated the identification of alkaline duodenogastric reflux and also eliminated its influence on the measurement of gastric bicarbonate secretion. Validation of the measuring system by instillation of small amounts of bicarbonate showed a satisfactory correlation between added and recovered bicarbonate in the range of bicarbonate determinations usually recorded. Decreasing intragastric pH to between 3 and 4 converted all secreted bicarbonate into CO2, but did not affect the measured value of bicarbonate secretion. Vagal stimulation accomplished by sham feeding increased gastric bicarbonate secretion in sixteen healthy subjects from 410 ± 39 µmol/h to 692 ± 67 µmol/h (mean ± SEM, p < 0.001). This response was independent of intragastric pH in the range of 2 to 7. The muscarinic receptor antagonist, benzilonium bromide, almost abolished the sham feeding response while indomethacin left it nearly unchanged. Nine duodenal ulcer patients had identical basal and vagally stimulated bicarbonate output as healthy subjects. Two months after proximal gastric vagotomy, the basal bicarbonate secretion was significantly increased, whereas the output in response to sham feeding was unaltered. In the early postoperative period, anticholinergics reduced the enhanced basal bicarbonate secretion to a preoperative level. In six healthy subjects, graded fundic distension with a balloon to volumes of 150 ml, 300 ml and 600ml, each distension period lasting 60 minutes, increased the bicarbonate secretion by 46 % (p < 0.05), 28 % (NS) and 84 % (p < 0.05), respectively. Continuous distension with 300 ml over 2.5 hours increased the bicarbonate secretion with a peak at 45 minutes, where after the response gradually declined. Seven duodenal ulcer patients investigated after proximal gastric vagotomy had a response to graded fundic distension virtually identical to that of healthy subjects. Anticholinergics abolished the response to fundic distension, whereas indomethacin was without any significant effect. In healthy subjects, 16,16_dimethyl PgE2 gave an about threefold greater response than vagal stimulation, fundic distension or carbachol. It is concluded that human gastric bicarbonate secretion is activated by cholinergic vagal nerves, the response being independent of intragastric pH at levels above pH 2. There seems to be an interplay of stimulatory and inhibitory mechanisms modulating basal as well as vagally stimulated gastric bicarbonate secretion. Fundic distension of the stomach stimulates bicarbonate secretion and the response is mediated by intramural neural cholinergic pathways. Neither vagal stimulation nor fundic distension apparently involves endogenous prostaglandin production. Vagal stimulation, fundic distension and carbachol appear to be submaximal stimuli of bicarbonate secretion or else may have both stimulatory and inhibitory actions on human gastric bicarbonate secretion. The increase in bicarbonate secretion during vagal stimulation and fundic distension parallels that of acid secretion and may be regarded as a physiological response to reinforce the protective mucus-bicarbonate barrier. I wish to express my sincere appreciation and gratitude to all colleagues, technicians, healthy volunteers and patients, who made this work possible. In particular, I want to thank my tutor, Lars Olbe, and those who helped me to carry out this thesis: Leif Hultén, Nils G. Kock, Gunnar Flemström, Lars Lundell, Bo Stenquist, Eva Albrektson, Gunilla Bogren, Eva-Lotta Eén, Marianne Moore, Barbro Nyman, Kerstin Olbe, Martin Riha and Karl Sandell. I want especially to thank Ingrid, my wife, Lisa and Gustav for their understanding and great support. The investigations were supported by grants from: AB Hässle, Mölndal, Sweden, Gothenburg Medical Society, Göteborg, Sweden, Glaxo Läkemedel AB, Mölndal, Sweden, Linmeko Design AB, Göteborg, Sweden, Medical Research Council of the Swedish Life Insurance
Companies, Stockholm, Sweden, Smith Kline & French AB, Solna, Sweden, Swedish Medical Research Council (project no. 17X-760), Stockholm, Sweden and Swedish Society for Medical Research, Stockholm, Sweden. Allen A. Structure and function of gastrointestinal mucus. In: Johnson LR, Christensen J, Grossman MI, Jacobson ED, Schultz SG,eds. Physiology of the gastrointestinal tract. Raven Press, New York1981:617-639. Allen A, Hutton D, McQueen S. Dimensions of gastroduodenal surface pH gradients exceed those of adherent mucus gel layers. Gastroenterology1983;85:463-466. Andre C, Bruhiere J, Vagne M, Lambert R. Bicarbonate secretion in the human stomach. Acta Hepato Gastroenterol 1973;20:62-69. Arvidsson S, Haglund U. Acute gastric ulceration in septic shock: an experimental model. Scand J Gastroenterol 1984;19 (suppl 105):46-48. Bahari HMM, Ross IN, Turnberg LA: Demonstration of a pH gradient across the mucus layer on the surface of human gastric mucosa in vitro. Gut 1982;23:513-516. Bell AE, Sellers LA, Allen A, Cunliffe WJ, Morris ER, Ross-Murphy SB. Properties of gastric and duodenal mucus: effect of proteolysis, disulfide reduction, bile, acid, ethanol, and hypertonicity on mucus gel structure. Gastroenterology 1985;88:269-280. Benos DJ. Amiloride: a molecular probe of sodium transport in tissues end cells. Am J Physiol 1982;242:C131-C145. Bickel M, Kauffman GL. Gastric gel mucus thickness: effect of distension,16,16-dimethyl prostaglandin E2 and carbenoxolone. Gastroenterology1981;80:770-775. Butler BD, Lichtenberger LM, Hills BA. Distribution of surfactants in the canine gastrointestinal tract and their ability to lubricate. Am J Physiol1983;244:G645-G651. Cheung LY, Chang N. The role of gastric mucosal blood flow and H back-diffusion in the pathogenesis of acute gastric erosions. J Surg Res1977;22:357-361. Cheung LY, Newton WT. Cyclic guanosine monophosphate response toacetylcholine stimulation of gastric alkaline secretion. Surgery1979;86:156-161. Crampton JR, Gibbons LC, Rees WDW. Effect of acid on bicarbonate secretion by the stomach in man: evidence for an autoregulatory reflex. Gut 1986;27(abstr):A592. Davenport HW. Gastric mucosal injury by fatty and acetylsalicylic acids. Gastroenterology 1964;46:245-253. Davenport HW. Fluid produced by the gastric mucosa during damage by acetic and acetylsalicylic acids. Gastroenterology 1966;50:487-499. Davenport HW. Salicylate damage to the gastric mucosal barrier. N Engl J Med 1976;276:1307-1312. Davenport HW. Back diffusion of acid through the gastric mucosa and its physiological consequences. In: Glass GBJ, eds. Progress in gastroenterology. Grune & Stratton, New York 1970;vol II:42-56. Debas HT, Walsh JH, Grossman MI. Evidence for oxyntopyloric reflex for release of antral gastrin. Gastroenterology 1975;68:687-690. Fahrenkrug J, Haglund U, Jodal M, Lundgren O, Olbe L, Schaffalitzky de Muckadell OB. Nervous release of vasoactive intestinal polypeptide in the gastrointestinal tract of cats: possible physiological implications. J Physiol 1978;284:291-305. Feldman M, Richardson CT, Taylor IL, Walsh JH. Effect of atropine on vagal release of gastrin and pancreatic polypeptide. J Clin Invest1979;63:294-298. Feldman M, Richardson CT. Gastric acid secretion in humans. In: Johnson LR, Christensen J, Grossman MI, Jacobson ED, Schultz SG, eds. Physiology of the gastrointestinal tract. Raven Press, New York 1981:693-708. Feldman M, Schiller L. Effect of bethanechol (Urecholine) on gastric acid and nonparietal secretion in normal subjects and duodenal ulcer patients. Gastroenterology 1982;83:262-266. Feldman M. Gastric bicarbonate secretion in humans. Effect of pentagastrin, bethanechol, and 11,16,trimethyl prostaglandin E2. J Clin Invest 1983;72:295-303. Feldman M, Colturi TJ. Effect of indomethacin on gastric acid and bicarbonate secretion in humans. Gastroenterology 1984;87:1339-1343. Feldman M. Gastric H+ and HCO3 secretion in response to sham feeding in humans. Am J Physiol 1985;11:G188-G191. Fich A, Arber N, Sestieri M, Zajicek G, Rachmilewitz D. Effect of misoprostol and cimetidine on gastric cell labeling index. Gastroenterology1985;89:57-61. Flemström G. Active alkalinization by amphibian gastric fundic mucosa in vitro. Am J Physiol 1977;233:E1-E12. Flemström G, Garner A. Stimulation of gastric acid and bicarbonate secretions by calcium in guinea pig stomach and amphibian isolated mucosa. Acta Physiol Scand 1980;110:419-426. Flemström G, Garner A. Gastroduodenal HCO3 transport: characteristics and proposed role in acidity regulation and mucosal protection. Am J 1982;242:G183-G193. Flemström G, Heylings JR, Garner A. Gastric and duodenal HCO3 transport in vitro: effects of hormones and local transmittors. Am J Physiol 1982;242:G100-G110. Flemström G, Turnberg LA. Gastroduodenal defence mechanisms. Clin Gastroenterol 1984;13:327-354. Flemström G. Measurement of gastric bicarbonate secretion. Gastroenterology 1985;88:2000-2002. Flemström G. Gastric and duodenal mucosal bicarbonate secretion. In: Johnson LR, Christensen J, Jackson MJ, Jacobson ED, Walsh JH, eds. Physiology of the gastrointestinal tract. Raven Press, New York 1987:1011-1029. Flemström G, Garner A. Secretion of bicarbonate by gastric and duodenal mucosa. In: Forte JG eds. Handbook of physiology. American Physiological Society, Bethesda. In Press. Forssell H, Preshaw R, Olbe L. Gastric secretion of bicarbonate determined with a perfusion technique. In: Allen A, Flemström G, Garner A, Silen W, Turnberg LA, eds. Mechanisms of mucosal protection in the upper gastrointestinal tract. Raven Press, New York 1984:125-128. I Forssell H, Olbe L. Continuous computerized determination of gastric bicarbonate secretion in man. Scand J Gastroenterol 1985;20:767-774. II Forssell H, Stenquist B, Olbe L. Vagal stimulation of human gastric bicarbonate secretion. Gastroenterology 1985;89:581-586. III Forssell H, Olbe L. Effect of proximal gastric vagotomy on basal and vagally stimulated gastric bicarbonate secretion in duodenal ulcer patients. Scand J Gastroenterol 1987;22:949-955. IV Forssell H, Olbe L. Effect of fundic distension on gastric bicarbonate secretion in man. Scand J Gastroenterol 1987;22:627-633. Furness JB, Costa M. The adrenergic innervation of the gastrointestinal tract. Rev Physiol 1974;69:1-51. Fändriks L, Delbro D. Neural stimulation of gastric bicarbonate secretion in the cat. An involvement of vagal axon-reflexes and substance P? Acta Physiol Scand 1983;118:301-304. Fändriks L. Sympatho-adrenergic inhibition of vagally induced gastroduodenal HCO3-secretion in the cat. Acta Physiol Scand 1986a;128:555-562. Fändriks L. Vagal and splanchnic neural influences on gastric and duodenal bicarbonate secretions. An experimental study in the cat. Acta Physiol Scand 1986b;128 (suppl 555):1-39. Gannon B, Browning J, O'Brien P, Rogers P. Mucosal microvascular architecture of the fundus and body of human stomach. Gastroenterology 1984;86:866-875. Gardham JR, Hoblsey M. The electrolytes of alkaline human gastric juice. Clin Sci 1970;39:77-87. Garner A, Flemström G. Gastric HCO3 secretion in the guinea pig. Am J Physiol 1978;234:E535-E541. Garner A, Heylings JR. Stimulation of alkaline secretion in amphibian-isolated gastric mucosa by 16,16-dimethyl PGE2 and PGF2: a proposed explanation for some of the cytoprotective actions of prostaglandins. Gastroenterology 1979;76:497-503. Garner A, Flemström G, Allen A. Gastroduodenal alkaline and mucus secretions. Scand J Gastroenterol 1983;18 (suppl 87):25-41. Gillespie IE, Clark DH, Kay AW, Tankel HI. Effect of antrectomy, vagotomy with gastrojejunostomy and antrectomy with vagotomy on the spontaneous and maximal gastric acid output in man. Gastroenterology 1960;38:361-367. Grötzinger U, Bergegĺrdh S, Olbe L. Effect of fundic distension on gastric acid secretion in man. Gut 1977a;18:105-110. Grötzinger U, Bergegĺrdh S, Olbe L. Effects of fundic distension on pentagastrin-stimulated gastric acid secretion in man. Gastroenterology1977b;73:447-452. Hasselbalch KA. Die Berechnung der Wasserstoffzahl des Blutes aus der freien und gebundenen Kohlensßäßure desselben,und die Sauerstoffbindung des Blutes als Funktion der Wasserstoffzahl. Biochem Z 1916;78:112-144. Hastings AB, Sendroy J. The effect of variation in ionic strength on the apparent first and second dissociation constants of carbonic acid. J Biol Chem 1925;65:445-455. Henderson LJ. The theory of neutrality regulation in the animal organism. Am J Physiol 1908;21:427-448. Heylings JR, Garner A, Flemström G. Regulation of gastroduodenal HCO3 transport by luminal acid in the frog in vitro. Am J Physiol 1984;246:G235-G242. Hills BA, Butler BD, Lichtenberger LM. Gastric mucosal barrier: hydrophobic lining to the lumen of the stomach. Am J Physiol 1983;244:G561-G568. Hills BA. Gastric mucosal barrier: stabilization of hydrophobic lining to the stomach by mucus. Am J Physiol 1985;249:G342-G349. Isenberg JI, Selling JA, Hogan DL, Koss MA. Impaired proximal duodenal mucosal bicarbonate secretion in patients with duodenal ulcer. N Engl J Med 1987;316:374-379. Ito S. Functional gastric morphology. In: Johnson LR, Christensen J,Grossman MI, Jacobson ED, Schultz SG, eds. Physiology of the gastrointestinal tract. Raven Press, New York 1981:517-550. Itoh M, Guth PH. Role of oxygen-derived free radicals in hemorrhagic shock-induced gastric lesions in rat. Gastroenterology 1985;88:1162-1167. Johansson C, Aly A, Nilsson E, Flemström G. Stimulation of gastric bicarbonate secretion by E2 prostaglandins in man. In: Samuelsson B, Paoletti R, Ramwell P, eds. Advances in prostaglandin, thromboxane and leukotriene research. Raven Press, New York 1983:395-401. Jönsson C, Fändriks L. Bleeding decreases duodenal HCO3 secretion by a nervous mechanism. Acta Physiol Scand 1986;127:273-274. Kauffman GL, Steinbach JH. Gastric bicarbonate secretion: effect of pH and topical 16-16 dimethyl prostaglandin E2 Surgery 1981;89:324-328. Kerss S, Allen A, Garner A. A simple method for measuring thickness of the mucus gel layer adherent to rat, frog and human gastric mucosa: influence of feeding, prostaglandin, N-acetylcysteine and other agents. Clin Sci1982;63:187-195. Kivilaakso E, Fromm D, Silen W. Relationship between ulceration and intramural pH of gastric mucosa during hemorrhagic shock. Surgery1978;84:70-78. Kivilaakso E, Fromm D, Silen W. Effect of the acid secretory state on intramural pH of rabbit gastric mucosa. Gastroenterology 1978;75: 641-648. Kivilaakso E. High plasma HCO3 protects gastric mucosa against acute ulceration in the rat. Gastroenterology 1981;81:921-927. Kivilaakso E. Contribution of ambient HCO3 to mucosal protection and intracellular pH in isolated amphibian gastric mucosa. Gastroenterology 1983;85:1284-1289. Kleist D, Bechtel H, Janisch HD, Bauer FE, Hampel KE. Effect of secretin on gastric pH and alkali secretion. Inn Med 1985;12:133-135. Konturek SJ, Bilski J, Tasler J, Laskiewicz J. Gastroduodenal alkaline response to acid and taurocholate in conscious dogs. Am J Physiol1984;247:G149-G154. Konturek SJ, Bilski J, Tasler J, Laskiewicz J. Gut hormones in stimulation of gastroduodenal alkaline secretion in conscious dogs. Am J Physiol1985;248:G687-G691. Lacy ER. Gastric mucosal resistance to a repeated ethanol insult. Scand J Gastroenterol 1985;20 (suppl 110):63-72. Lichtenberger LM, Graziani LA, Dial EJ, Butler BD, Hills BA. Role of surface-active phospholipids in gastric cytoprotection. Science1983;219:1327-1329. Lichtenberger LM, Richards JE, Hills BA. Effect of 16,16-dimethylprostaglandin E2 on the surface hydrophobicity of aspirin-treated canine gastric mucosa. Gastroenterology 1985;88:308-314. Lind T, Cederberg C, Axelson M, Olbe L. Long-term acid inhibitory effect on different daily doses of omeprazole 24 hours after dosing. Scand J Gastroenterol 1986;21 (suppl 118):137-138. Lönnerholm G, Selking Ö, Wistrand PJ. Amount and distribution of carbonic anhydrases CA I and CA II in the gastrointestinal tract. Gastroenterology 1985;88:1151-1161. Martinson J, Muren A. Excitatory and inhibitory effects of vagus stimulation on gastric motility in the cat. Acta Physiol Scand 1963;57:309-316. Noring O. Studies on the cephalic phase of gastric secretion in normal subjects and ulcer patients. Gastroenterology 1951;18:413-418. Nylander O, Flemström G, Delbro D, Fändriks L. Vagal influence on gastroduodenal HCO3-secretion in the cat in vivo. Am J Physiol 1987 in press. Odes HS, Hogan DL, Koss MA, Ballesteros MA, Steinbach JH, Isenberg JI. Methods used to measure gastric bicarbonate secretion in humans: why do they result in different values? Gastroenterology 1987;92:no.5(2), abstract. Powel DW. Barrier function of epithelia. Am J Physiol 1981;241: G275-G288. Quigley EMM, Turnberg LA. Identification in vivo of a neutral pH zone lining gastroduodenal mucosa in man. Dig Dis Sci 1985;30:396. Rees WDW, Botham D, Turnberg LA. A demonstration of bicarbonate production by the normal human stomach in vivo. Dig Dis Sci1982;27:961-966. Reinhart WH, Müller O, Halter F. Influence of long-term 16,16-dimethyl prostaglandin E2 treatment on the rat gastrointestinal mucosa. Gastroenterology 1983;85:1003-1010. Ritchie WP Jr, Shearburn EW III. Influence of isoproterenol and cholestyramine on acute gastric mucosal ulcerogenesis. Gastroenterology1977;73:62-65. Ritchie WP Jr. Acute gastric mucosal damage induced by bile salts, acid, and ischemia. Gastroenterology 1975;68:699-707. Ross IN, Bahari HMM, Turnberg LA. The pH gradient across mucus adherent to rat fundic mucosa in vivo and the effect of potential damaging agents. Gastroenterology 1981;81:713-718. Rune SJ. The metabolic alkalosis following aspiration of gastric secretion. Scand J Clin Lab 1965;17:305_310. Rune SJ, Henriksen FW. Carbon dioxide tensions in the proximal part of the canine gastrointestinal tract. Gastroenterology 1969;56:758-762. Rutten MJ, Ito S. Morphology and electrophysiology of guinea pig gastric mucosal repair in vitro. Am J Physiol 1983;244:G171-G182. Schierbeck NP. Ueber Kohlensßäßure im Ventrikel. Scand Arch Physiol 1892;3:437-473. Schöön I-M, Bergegĺrdh S, Grötzinger U, Olbe L. Evidence for a defective inhibition of pentagastrin-stimulated gastric acid secretion by antral distension in the duodenal ulcer patient. Gastroenterology 1978;75:363-367. Selling JA, Hogan DL, Aly A, Koss MA, Isenberg JI. Indomethacin inhibits duodenal mucosal bicarbonate secretion and endogenous PGE2 output in humans. Ann Internal Med 1987;106:368_371. Skov Olsen P, Poulsen SS, Kirkegaard P, Nexo E. Role of submandibular saliva and epidermal growth factor in gastric cytoprotection. Gastroenterology 1984;87:103-108. Slomiany BL, Slomiany A. Glycosphingolipids, glyceroglucolipids of glandular epithelial tissue. In: Sweeley CC, eds. Cell surface glycolipids. Am Chem Soc, Washington 1980:149-176. Slyke van DD, Sendroy J, Hastings AB, Neill JM. Studies on gas and electrolyte equilibria in blood. The solubility of carbon dioxide at 38° in water, salt solution, serum and blood cells. J Biol Chem 1928;78:765-799. Smith GW, Tasman-Jones C, Wiggins PM, Lee SP. Pig gastric mucus: A one-way barrier for H+. Gastroenterology 1985;89:1313-1318. Stenquist B, Knutsson U, Olbe L. Gastric acid response to adequate and modified sham feeding and to insulin hypoglycemia in duodenal ulcer patients. Scand J Gastroenterol 1978;13:357-362. Stenquist B, Rehfeld JF, Olbe L. Effect of proximal gastric vagotomy and anticholinergics on the acid and gastrin responses to sham feeding in duodenal ulcer patients. Gut 1979;20:1020-1027. Stenquist B, Lind T, Haglund U, Holst JJ, Rehfeld JF, Olbe L. Do enkephalins participate in vagal activation of gastric acid secretion in man? Regul Peptide 1987;17:1_7. Sugai N, Ito S. Carbonic anhydrase, ultrastructural localization in the mouse gastric mucosa and improvements in the technique. J Histochem Cytochem1980;6:511-525. Sung CP, Wiebelhaus VD, Jenkins BC, Adlercreutz P, Hirschowitz BI, Sachs G. Heterogenity of 3',5'-phosphodiesterase of gastric mucosa. Am J Physiol 1972;223:648-650. Svanes K, Ito S, Takeuchi K, Silen W. Restitution of the surface epithelium of the in vitro frog gastric mucosa after damage with hyperosmolar sodium chloride. Gastroenterology 1982;82:1409-1426. Svanes K, Takeuchi K, Ito S, Silen W. Effect of luminal pH and nutrient bicarbonate concentration on restitution after gastric surface injury. Surgery 1983;94:494-500. Svanes K, Critchlow J, Takeuchi K, Magee D, Ito S, Silen W. Factors influencing reconstitution of frog gastric mucosa: role of prostaglandins. In: Allen A, Flemström F, Garner A, Silen W, Turnberg LA, eds. Mechanisms of mucosal protection in the upper gastrointestinal tract. Raven Press, New York 1984:33-38. Szabo S, Trier JS, Frankel PW. Sulfhydryl compounds may mediate gastric cytoprotection. Science 1981;214:20-22. Szabo S. Peptides, sulfhydryls, and glucocorticoids in the gastric mucosal defense: coincidence or connection? Gastroenterology 1984;87:228-229. Säfsten B, Flemström G. Stimulatory effect of pirenzipine on mucosal bicarbonate secretion in rat duodenum in vivo. Acta Physiol Scand 1986;127:267-268. Takeuchi K, Magee D, Critchlow J, Matthews J, Silen W. Studies of the pH gradient and thickness of frog gastric mucus gel. Gastroenterology1983;84:331-340. Takeuchi K, Tanaka H, Furukawa O, Okabe S. Gastroduodenal HCO3 secretion in anesthetized rats: effects of 16,16-dimethyl PgE2, topical acid and acetazolamide. Japan J Pharmacol 1986;41:87-99. Tasman-Jones C. Gastric mucus - physical properties in cytoprotection. Med J Austr 1985;142 (suppl):S5-S6. Teorell T. On the stomach permeability for acids and some other substances.J Gen Physiol 1939;23:263-274. Tytgat GNJ, Offerhaus GJA, Van Minnen AJ, Everts V, Hensen-Logmans SC, Samson G. Influence of oral 15(R)-15-methyl prostaglandin E2 on human gastric mucosa. A light microscopic, cell kinetic, and ultrastructural study. Gastroenterology 1986;90:1111-1120. Wallace JL, Whittle BJR. Role of mucus in the repair of gastric epithelial damage in the rat. Inhibition of epithelial recovery by mucolytic agents. Gastroenterology 1986;91:603-611. Werther JL, Hollander F, Altamirano M. Effect of acetazolamide on gastric mucosa in canine vivo-vitro preparations. Am J Physiol 1965;209:127-133. Williams SE, Turnberg LA. Retardation of acid diffusion by pig gastric mucus: a potential role in mucosal protection. Gastroenterology1980;79:299-304. Williams SE, Turnberg LA. Demonstration of a pH gradient across mucus adherent to rabbit gastric mucosa: evidence for a 'mucus-bicarbonate barrier. Gut 1981;22:94-96. Wright NA. Role of mucosal cell renewal in mucosal protection in the gastrointestinal tract. In: Allen A, Flemström G, Garner A, Silen W, Turneberg LA, eds. Mechanisms of mucosal protection in the upper gastrointestinal tract. Raven Press, New York 1984;:15-19. Younes M, Schoenberg MH, Jung H, Fredholm BB, Haglund U, Schildberg FW. Oxidative tissue damage following regional intestinal ischemia and reperfusion in the cat. Res Exp Med 1984;184:259-264. Zalewsky CA, Moody FG. Mechanisms of mucus release in exposed canine gastricmucosa. Gastroenterology 1979;77:719-729. |